ЭВОЛЮЦИЯ ЖИЗНИ
– ПРОИЗВОДНАЯ РАЗВИТИЯ ГАЗОВОГО РЕЖИМА НА ПЛАНЕТЕ ЗЕМЛЯ.
15.01.2006г.
Введение
Планета Земля,
как составная часть солнечной системы, сформировалась (по современным
представлениям) примерно 4 – 4.5 млрд. лет назад. О газовом составе
протоатмосферы мы можем судить только по косвенным признакам. Одними из
основных газов являлись азот, углекислый газ, пары воды. Также присутствовали
другие газы и вещества в газообразном состоянии, устойчивые для тех
термодинамических условий, которые господствовали на поверхности молодой
планете Земля. В результате остывания планеты была достигнута
точка росы, т.е. температура начала конденсации паров воды, для тех
термодинамических условий земной атмосферы. На поверхность Земли “устремились”
потоки сконденсировавшейся воды, которые дали начало морям и океанам. Земля
обрела ещё одну оболочку – гидросферу. Ювенильная вода, поступающая из недр
Земли, теперь уже не только испарялась в атмосферу, но и пополняла объём
гидросферы.
Одно можно
смело утверждать, что свободного кислорода в атмосфере молодой Земли не было.
Неоспоримым подтверждением этого утверждения является наличие окатанного пирита
в осадочных породах протерозойского месторождения Витватерсранд, на юге Африки.
Пирит (FeS2)
является неустойчивым минералом в окислительной обстановке, быстро разлагается
с образованием лимонита и солей серной кислоты, которые, как правило выносятся
водой за пределы коры выветривания. Следовательно, пирит можно использовать как
индикатор восстановительной и окислительной обстановки. Присутствие окатанного
пирита в осадочных горных породах, говорит только о его механической обработке
в экзогенных процессах и отсутствии химического разложения. Отсюда вывод – протерозойская атмосфера была
бескислородной.
Архейская атмосфера, как более древняя, также была бескислородной. Нет свободного кислорода – нет и озонового слоя, поэтому, жизнь на базе углерода была возможна только в водной среде. Ультрафиолетовое излучение молодого Солнца уничтожало всякие зародыши жизни на поверхности Земли вне водной среды.
Вопрос о происхождение жизни на планете
Земля не входит в круг рассмотрения предлагаемой читателю гипотезы. Автор лишь
увязывает снижение содержания углекислого газа и параллельное повышение
содержания кислорода в морской воде и атмосфере Земли, с вымиранием одних и
появлением других форм флоры и фауны.
Древнейшие ископаемые остатки флоры, синезелёные водоросли, имеют возраст более 3 млрд. лет. Отсюда следует, что в результате фотосинтеза, процесс поступления свободного кислорода в морскую воду и далее в атмосферу начался ещё в архейской эре. Первоначально выделявшийся кислород расходовался на перевод закисных форм химических элементов в окисные, т.е. на преобразование восстановительной обстановки на поверхности планеты, в окислительную. Следовательно, аэробная фауна появилась значительно позже флоры, когда количество свободного кислорода в среде обитания (в морской воде) стало достаточным для дыхания животных организмов.
ФАКТОРЫ, ВЗЫВАЮЩИЕ КАЧЕСТВЕННЫЕ СКАЧКИ
В ИСТОРИИ РАЗВИТИЯ ЖИЗНИ НА ПЛАНЕТЕ ЗЕМЛЯ
Флора
Одним из необходимых условий для жизни растений является наличие углекислого газа. В первичной атмосфере углекислый газ присутствовал в избытке. Первые растения были приспособлены именно к большому количеству углекислого газа в атмосфере. Но сами же растения, в результате фотосинтеза, начали понижать содержание углекислого газа в воде с последующим его уменьшением и в атмосфере. Увеличивающаяся светимость молодой звезды, Солнца, способствовала увеличению скорости фотосинтеза на планете Земля.
Изъятие углекислого газа из морской воды происходило не только в результате фотосинтеза. Первоначально часть синезелёных водорослей, а в дальнейшем, с появлением фауны, и многие представители животных, активно поглощали СО2 (спикулы губок, раковинки состоящие из СаСО3 и др.). Поставщиками кальция были коры выветривания, которые развивались по кальций содержащим горным породам типа габбро, базальта, диорита, андезита и д.т. В результате на планете Земля образовалось огромное количество органогенных известняков (стромотолитовый известняк, известняк ракушечник, мел, коралловый известняк и др.). Основная часть мрамора также образовалась в результате метаморфизма органогенных известняков.
Изымание углекислого газа из морской воды происходило и происходит также в результате химических процессов, протекающих в водной среде. Это подтверждается наличием в земной коре значительного количества хемогенных известняков самого разного возраста среди осадочных горных пород.
Снижение содержания углекислого газа способствовало и понижение температуры на поверхности Земли, т.к. с понижением температуры растворимость газов, в том числе и СО2, в жидкой воде увеличивается, а образование СаСО3 происходит в основном в водной среде.
К снижению содержания углекислого газа в воде и атмосфере ведёт и вывод части углерода из природного круговорота в виде углеводородных газов, торфа, бурого угля, органического детрита, которого достаточно много во многих осадочных горных породах. Каменный уголь и нефть также вывели из круговорота огромное количество углерода (если они действительно органического происхождения).
Понижение содержания углекислого газа в среде обитания являлось причиной эволюции самих растениях.
Один из главных философских законов гласит: количественные изменения приводят к изменению качества. Из этого закона вытекает и такое понятие как мера.
Мерой
называется количество, определяющее переход в качество, т.е. превышение меры, в
сторону уменьшения или увеличения количества, ведёт к скачкообразному изменению
качества.
В результате выше описанных процессов на планете Земля содержание углекислого газа непрерывно снижалось. Количественная мера, вызывающая качественные скачки в эволюции растительного мира, в сторону уменьшения СО2, превышалась много раз. В нашем случае удобней говорить о достижении порогового содержания СО2, что не меняет сути, после чего начиналось быстрое вымирание большинства старых и появление новых растений, более приспособленных к снизившемуся содержанию углекислого газа в окружающей среде. Часть старых растений приспосабливалась к новым условиям, занимая мало благоприятные экологические ниши. Но свои позиции они уступали более приспособленным к изменившемуся содержанию СО2 новым формам растений. После чего наступал период ”спокойной“, дарвиновской, эволюции до достижения следующего порогового содержания СО2 в биосфере Земли.
Таким образом, уменьшение содержания СО2 в воде и атмосфере принуждало флору постоянно скачкообразно обновляться. В результате флорой, в своём развитии, был пройден путь от низших растений до высших (синезелёные водоросли – покрытосеменные растения). На этом пути скачкообразные изменения происходили достаточно часто (в геологическом понимании). Например, пермская смена споровых растений голосеменными растениями, которые господствовали потом на суше планеты почти всю мезозойскую эру. В свою очередь голосеменные растения, в меловом периоде, сменились покрытосеменными растениями. Основная часть голосеменных растений вымерла, а оставшаяся находится в подчинённом состоянии по отношению к покрытосеменным растениям.
Поставщиками углекислого газа на поверхность Земли на начальном этапе были вулканы, магматические интрузии, глубинные разломы в земной коре т.е. её недра. С появлением свободного О2 и фауны на планете, СО2 стал выделяться в результате дыхания живых организмов, а также в результате сгорания органического вещества. Однако снижение содержания СО2 на поверхности Земли преобладало над его поступлением, а поступление кислорода преобладало над его расходованием. Конечный результат – это нынешнее содержание СО2 (~0.04%) и О2 (~20%) в атмосфере. В настоящее время СО2 поступает, а О2 изымается из атмосферы в результате активной деятельности человека, что нарушает природный баланс изъятия и поступления СО2 и О2 в биосферу. Это может привести к непредсказуемым последствиям.
Эпохи интенсивного выделения СО2 с незначительными задержками во времени сопровождались эпохами буйства растительной жизни. Если верить в органическое происхождение основной части существующих на планете углей, то именно расцвет флоры в каменноугольном периоде предопределил смену споровых растений на голосеменных в пермском периоде. Снижение содержания в морской воде и в атмосфере СО2 сопровождалось повышением содержания О2, что привело к упадку земноводных и появлению в триасе рептилий (динозавров в том числе) и примитивных млекопитающих.
Достижение и
превышение порогового содержания СО2, в сторону уменьшения,
сопровождалось вымиранием одних и появлением других форм растительности
повсеместно на всей планете. Появлялись не только новые виды, но и новые типы растений.
Это объясняется тем, что воздух в атмосфере и вода в океане достаточно быстро перемешиваются, что ведёт к повсеместному выравниванию содержания СО2 в биосфере, с учётом зональности его распределения в воде и атмосфере.
Отсюда следует, что при достижении порогового
содержания СО2,в сторону уменьшения, вымирание происходило по всей
Земле, независимо от широты, климата, высоты над уровнем океана и места
произрастания растений (в воде или на суше).
Вот почему на лике Земли вроде бы неизвестно откуда появлялись ранее не существовавшие, совершенно новые формы жизни, и без видимой причины исчезали ранее процветавшие формы.
Именно такие смены мы довольно часто наблюдаем в истории развития флоры и фауны на планете Земля.
Фауна
Животный мир на планете Земля во времени появился несколько позднее растительного. Возникновению животных предшествовало появление свободного кислорода в морской воде. В кембрийском периоде уже жили и процветали достаточно высокоорганизованные животные типа медуз, трилобитов, ракоскорпионов, плеченогих и других. Внутри кембрийского животного мира уже существовало разделение на растительноядных и плотоядных животных. Это говорит о том, что появление примитивной фауны произошло значительно раньше. Первые животные на Земле появились и жили при низком содержании кислорода в морской воде. Атмосфера всё ещё оставалась практически бескислородной. В результате развития флоры, как в количественном, так и в качественном направлении, интенсивность фотосинтеза непрерывно повышалась. Следовательно, содержание свободного кислорода в морской воде и атмосфере непрерывно увеличивалось, а содержание углекислого газа уменьшалось (Рис.1). Эти изменения были непропорциональными, т.к. на синтез флорой органического вещества с выделением О2, расходовалась лишь часть СО2. Чтобы кислород накапливался в атмосфере и в океанической воде Земли необходимо удаление части углерода из природного круговорота. Удаление углерода из природного круговорота происходит при осадконакоплении в виде детрита из органических остатков, торфа, бурого угля, углеводородного газа и т.д. Нынешнее вовлечение захороненного углерода в природный круговорот может понизить содержание О2 что приведёт к катастрофическим последствиям
С появлением свободного кислорода в атмосфере начал
формироваться озоновый слой. Это позволило жизни начать освоение новой среды
обитания – суши. Из животных, одними из первых на сушу вышли насекомые, при ещё
низком содержании кислорода в атмосфере.
Схематические графики
изменения содержания СО2 и О2
в истории Земли
Содержание СО2
![]() и О2 в атмосфере (%)
и О2 в атмосфере (%)
![]() 100
100
Содержание СО2 в атмосфере
Фанерозой![]()
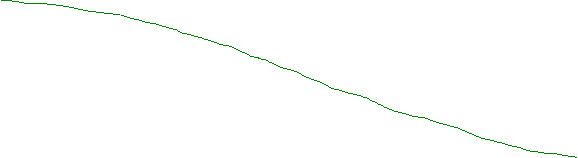
(PZ+MZ+KZ)
Криптозой
(докембрий)
![]() 25
25

![]()
![]()
![]() 0.4
0.4
3.43 4 Тмлрд. лет
Рис.1
Предлагаемая на суд читателю гипотеза
основывается на том, что в каждый исторический промежуток времени основная
часть фауны была приспособлена к строго определённому содержанию кислорода в
окружающей среде. Даже относительно незначительные колебания содержания
кислорода приводили к вымиранию одних и появлению других таксонометрических
единиц животных.
Различия в
животном мире между соседними ярусами одного и того же периода незначительные, но всё-таки
существуют.
При постоянном
содержании кислорода в воде и атмосфере эволюция фауны идёт по дарвиновскому
сценарию, т.е. могу появляться в только новые виды, подвиды, но не более того.
Увеличение свободного О2, достигнув порогового содержания в окружающей среде, угнетающе действовало на животный мир и в течение короткого времени (по геологическим меркам) многие, господствовавшие ранее таксонометрические единицы животных вымирали. Выживала лишь незначительная часть животных старого мира, приспособившись к повышенному содержанию О2, оставаясь теперь на второстепенных ролях. Иногда старые формы животных занимали незанятые экологические ниши, близкие к их старым условиям. Так поступили морские лилии, кистепёрые рыбы, шестижаберные акулы и др.. В своём расцвете морские лилии жили в мелких морях конца палеозоя, но с увеличением содержания кислорода в приповерхностных слоях морской воды, ушли на глубины до 10км, куда не проникают солнечные лучи, не растут водоросли, а следовательно и содержание О2 несколько ниже.
Следует
отметить, что революционные изменения в составе флоры всегда шли с отставанием
от таких же изменений в фауне. Особенно наглядно это видно при смене
мезозойской флоры и фауны кайнозойской. Смена голосеменных растений
покрытосеменными произошла в меловом периоде. Следовательно, к началу мелового
периода было достигнуто пороговое содержание СО2, что привело к
революционным изменениям в растительном мире. А вот пороговое содержание
кислорода было достигнуто лишь к началу кайнозойской эры, что привело, к
вымиранию всех динозавров, как сухопутных, так и морских, а также и других
классов и семейств позвоночных животных. Вымирание коснулось и непозвоночных
животных. Так вымерли почти все головоногие моллюски
– аммоноидеи. Сумел приспособиться к новому, более высокому содержанию
кислорода в морской воде лишь их один вид – наутилус, да и то он находится в
угнетённом состоянии (индивидуумы небольших размеров, практически нет
скульптуры на раковинках, не чета мезозойским аммонитам).
В истории фауны имели место и случаи, когда повышение О2 было благоприятным для расцвета примитивных животных, например расцвет нуммулитов в кайнозое.
Даже незначительное увеличение содержания кислорода вызывало смену одних сообществ животных другими. Эти изменения уже видны между фауной периодов и ярусов. Так кембрийские трилобиты довольно сильно отличаются от ордовикских и тем более от силурийских, а в девоне они исчезли вообще.
Наблюдается закономерность, чем выше
становилось содержание кислорода в окружающей среде, тем более сложнее
устроенные и более активные появлялись типы, классы, семейства, роды и виды
животных.
Наглядно это подтверждается эволюцией хордовых. Эволюция рыб от бесчелюстных, панцирных, до костных, а также эволюция наземных животных (земноводные → рептилии → плацентарные млекопитающие). Первые рептилии (динозавры в том числе) появились на Земле практически одновременно (по геологическим меркам) с первыми яйцекладущими млекопитающими, это примерно середина и конец триасового периода соответственно. Содержание кислорода в начале, да и в течение всей мезозойской эры для расцвета млекопитающих было недостаточно, поэтому они до конца мезозойской эры были в тени у динозавров, “влача жалкое существование”. Для динозавров же наоборот, содержание кислорода в триасовом, юрском и меловом периодах оказалось оптимальным, т.е. содержания кислорода от триасового периода до палеогенового хоть и повышалось, но оставалось для них в пределах меры.
Этим объясняется расцвет динозавров в мезозойскую эру. Но динозавры и аммониты, да и многие другие животные, оказались стеногалинными животными по отношению к содержанию кислорода в атмосфере и в морской воде. На границе между мезозойской и кайнозойской эрами динозавры и их современники просто “сгорели”, каждый в своей среде обитания, так как количество кислорода для них в воде и атмосфере превысило пороговое содержание. Повышенное содержание кислорода оказалось губительным практически для всех яйцекладущих и подавляющего большинства сумчатых млекопитающих, которые последовали за своими современниками – динозаврами.
Содержание кислорода в атмосфере ~18–20% можно
считать губительным для всех динозавров и оптимальным для птиц и плацентарных
млекопитающих.
История появления и расцвета птиц, также вписывается в предлагаемую читателю гипотезу. Почему на планете Земля, согласно данным палеонтологии, так поздно появились птицы? Для жизнедеятельности птиц необходимо бÓльшее содержание кислорода в атмосфере, чем для динозавров и примитивных млекопитающих, из-за более активного обмена веществ. Это связано с поддержанием высокой (~42 оС) температуры тела. Вот почему появление птиц на Земле задержалось на десятки миллионов лет, и лишь с достижением для них порогового содержания кислорода в атмосфере юрского периода, стало возможным появление нового класса хордовых – класса птиц. Первые птицы, из-за ещё низкого для них в юрском периоде содержания кислорода, были планирующими, т.к. планирующий полёт требует меньшей затраты энергии.
Практически все классы животных в своём развитии переживали
4 эпохи:
1.
Появление
примитивных форм животных (содержание кислорода минимально достаточное для их
существования). Млекопитающие – триас, птицы – верхняя юра и т.д.
2.
Гигантизм хорошо
сформированных, развитых форм животных (содержание кислорода оптимальное). Рыбы
– девон; насекомые – карбон; стегоцефалы –
конец карбона; динозавры– юра, плацентарные млекопитающие –неоген.
3.
Угнетённые формы
животных (содержание кислорода максимально допустимое для их
жизнедеятельности). Стегоцефалы – пермь; динозавры – меловой период.
4.
Вымирающие формы
животных (содержание кислорода превысило предельно допустимое содержание для
существующих форм животных. Оно же вызовет появление новых, более
приспособленных к изменившемуся содержанию О2 , форм животных).
Это относится и к флоре.
![]() Таким
образом, чем выше содержание кислорода в среде обитания, тем более
высокоорганизованные растения и животные занимают в ней господствующее положение
(рыбы → земноводные → рептилии → плацентарные млекопитающие и
птицы).
Таким
образом, чем выше содержание кислорода в среде обитания, тем более
высокоорганизованные растения и животные занимают в ней господствующее положение
(рыбы → земноводные → рептилии → плацентарные млекопитающие и
птицы).
Эту закономерность можно проследить и среди морских беспозвоночных животных, но наиболее ярко она видна для хордовых представителей фауны.
На размеры фауны и флоры могут влиять эпохи сжатия и расширения планеты, которые ведут к изменению силы тяжести на поверхности Земли. При увеличении радиуса планеты сила тяжести на её поверхности уменьшается и наоборот. Следовательно, для выяснения причин гигантизма следует применять комплексный подход.
Изменения СО2 и О2
вынуждают фауну и флору
совершать скачки в своём развитии
Согласно законам генетики, любой
вид живых организмов сопротивляется любому изменению в его ДНК. Опыты с
бактериями показывают, что размножение путём простого деления преобладает в том
случае, если они живут в благоприятной среде. Поместив этих же бактерий в
неблагоприятную среду, способ размножения у них резко изменяется. Теперь часто,
перед делением две бактерии сливаются вместе, и только потом происходит
деление. Слияние происходит не хаотично, а выборочно. Две бактерии сливаются
вместе лишь в том случае, если у них повреждены разные участки ДНК или одна из
них вообще их не имеет.
После деления объединившейся бактерии обе новые бактерии имеют целые,
неповреждённые ДНК, т.е. происходит замена повреждённых участков ДНК
неповреждёнными.
Так работает защита, препятствующая какому-либо изменению вида. Возможно, это является причиной появления двуполого способа размножения у многих представителей флоры и фауны на планете Земля.
При достижении пороговых
содержаний СО2 или О2 в флоре или фауне соответственно, происходит
падение практически до нуля защиты от каких-либо генетических изменений (идёт
процесс вымирания), а перед этим организмы жили в неблагоприятной газовой
среде. Появляется огромное количество мутантов. Как правило, основная часть их
не жизнеспособна, но встречаются и приспособленные к изменившимся условиям (по
газовому составу) среды обитания. Далее, новые представители флоры и фауны
начинают быстро занимать освободившиеся экологические ниши, переводя выживших
представителей старых экосистем на второстепенные роли.
Революционные изменения между эрами это изменения первого порядка. Внутри пороговых содержаний первого порядка существуют пороговые содержания второго, третьего и т.д. порядков. В зависимости от того, какого порядка достигнут порог содержания СО2 или О2 и происходит смена видов или родов, или семейств и т.д. представителей флоры и фауны. В истории жизни это наблюдается как смена флоры и фауны между эрами, периодами, эпохами, ярусами.
Отсюда вытекает вывод, что стратиграфическое расчленение возраста горных пород по палеонтологическим данным, это нечто иное, как вехи в истории развития газового режима на поверхности планеты Земля.
Изменится ли содержание кислорода в будущем?
За счёт нынешнего содержания углекислого газа в атмосфере и гидросфере значительное повышение кислорода в атмосфере уже не возможно. Остаются два источника, это космос и земные недра. Из космоса поступление СО2 возможно но мало вероятно. А то, что в недрах Земли присутствуют кислород и углерод подтверждают вулканы (обилие СО2 среди вулканических газов), карбонатитовые интрузии, ореолы карбонатной гидротермальной минерализации вокруг гранитных и других по составу магматических пород. Земля является геологически активной планетой, следовательно, резкий выброс углекислого газа в больших объёмах особенно в эпоху временного планетарного сжатия вполне возможен. А дальше уже по накатанному пути, резкое увеличение биомассы растений с быстрым повышением концентрации кислорода в атмосфере со всеми вытекающими отсюда последствиями.
Выживут ли плацентарные животные, если содержание
кислорода в атмосфере поднимется до 25-30%? Вряд ли, но кто займёт их место? И не вернутся ли динозавры на Землю, если
содержание О2 в атмосфере опустится до мезозойского, и что станет с
современным человеком?
В настоящее время всё чаще и чаще рождаются
дети с неординарными способностями. Не началась ли уже замена нынешнего Homo sapiens на его
новый вид?
На раннем этапе развития светимость молодой звезды, Солнца, увеличивалось, что вело к увеличению скорости фотосинтеза на планете Земля. В будущем, когда светимость Солнца начнёт уменьшаться, начнёт уменьшаться и скорость фотосинтеза, что приведёт к увеличению СО2 и понижению О2 и тогда эволюция жизни на планете Земля, может повернуться вспять.
Влияние эпох орогенеза на флору и фауну
Эпохи орогенеза (вернее эпохи временного сжатия расширяющейся планеты) всегда сопровождаются интенсивным вулканизмом центрального типа, мощными землетрясениями, приводящими к сокращению площади земной коры (это спрединг, надвиги, шарьяжи и др.). Внедрением в земную кору магматических интрузий, а также полное отсутствие на планете активности вулканизма трещинного типа (в настоящее время (2006г.) всё это имеет место). Все эти геологические процессы приводят к выбросу на поверхность Земли (в атмосферу и в морскую воду) огромного количества углекислого газа. С увеличением СО2 увеличивалась биомасса растений. Но при особо интенсивных эпохах сжатия планеты (например: калидонская, герцинская, киммерийская эпохи орогенеза, а вернее эпохи сжатия и др.) возможно выделялось такое огромное количество углекислого газа, что при его переработке растениями, могло резко повышаться содержание кислорода, которое приводило к выше описанным процессам. Увеличению содержания свободного кислорода способствует вывод части углерода из круговорота в виде растительного детрита (присутствует во многих осадочных горных породах), торфа, бурого угля, а также в виде каменного угля и нефти, если верить в их органическое происхождение.
Заключение
В выше изложенной статье затронута лишь незначительная часть процессов, вызванных изменением содержания СО2 и О2 в атмосфере планеты Земля.
В статье не затронуты революционные перестройки жизни на границе между рифеем, вендом и палеозоем. Пути изменения форм жизни между периодами в пределах отдельно взятых эр. Весьма интересно с новых позиций проследить путь развития насекомых (гигантизм муравьёв, стрекоз, тараканов и др. в карбоне).
Весьма интересно было бы знать количественные содержания СО2 и О2 для отдельно взятых эр, периодов и эпох. Ведь тогда появляется возможность вернуть вымерших животных или получить совершенно новые формы живых существ, не рискуя быть уничтоженными продуктами генной инженерии (бесплодные свиньи, которых кормили генетически изменённым картофелем).